Seabed Habitats
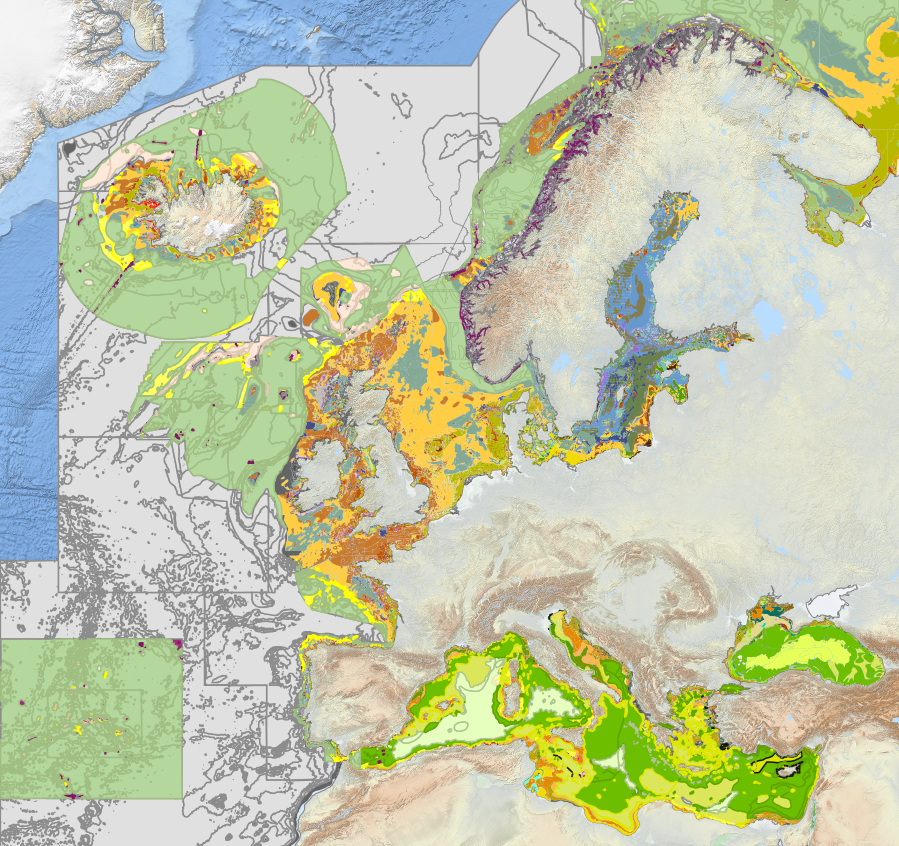
Seabed Habitats provides access to seabed habitat data in Europe. This includes new products such as the EMODnet broad-scale seabed habitat map for Europe (AKA EUSeaMap) and habitat maps and observations gathered from surveys across Europe.
Background
Since its inception in 2009, EMODnet Seabed Habitats has developed, improved and gradually increased the coverage of a broad-scale seabed habitat map Europe's seabed, also known as EUSeaMap. The most recent iteration was created in 2021 using the latest seabed substrate from EMODnet Geology.
In addition, EMODnet Seabed Habitats continued the work started by MESH and MESH Atlantic projects in collating and making available seabed habitat maps from surveys, through the map viewer.
In its third Phase (2017-2021), EMODnet Seabed Habitats began to collate and provide habitat point data and the outputs of habitat distribution modelling. In addition, this phase saw the creation of new composite data products, which combine data from the existing collation of habitat maps and observations to show the distribution of specific habitats. Examples in the the Essential Ocean Variable habitats Live hard coral cover, Seagrass cover and Macroalgal canopy cover.
Phase four (2021-2023). In this phase EMODnet Seabed Habitats will:
- Continue to grow Europe's only comprehensive library of habitat maps from surveys and collection of survey sample points;
- Expand its remit to coastal wetlands and essential fish habitats;
- Create new composite data products showing the extent and distribution of important habitats in and around Europe;
- Update the EMODnet broad-scale seabed habitat map for Europe (EUSeaMap), including two new regions: the Caspian Sea and some European territories in the Caribbean.
EUSeaMap - Broad-scale Seabed Habitat Map for Europe
The EMODnet broad-scale seabed habitat map for Europe is a comprehensive, free and ready-to-use broad-scale map of physical habitats, harmonising mapping procedures and fostering a common understanding among seabed mappers in Europe. The map is also known as EUSeaMap.
The EUSeaMap broad-scale predictive mapping methods are repeatable and ensure that the predictive maps can continue to be improved in the future. EUSeaMap is available in the EUNIS and MSFD Benthic Broad Habitat types classifications.
Building on the highly successful INTERREG IIIB-funded MESH and BALANCE projects, phase I of EMODnet Seabed Habitats (2009-2012) improved and harmonised the broad-scale physical habitat map across the Celtic Seas, Greater North Sea and Baltic Sea, as well as generating the very first broad-scale map of the western Mediterranean. In phase II (2013-2016), the coverage of the maps was extended to all European seas and the existing maps were improved. Updates and further increases in the extent to include the Barents Sea took place in phase III (2017-2019).
The data layers and confidence maps produced by project partners, are freely available through the Map Viewer. More information on the methodology used to create the most recent version of EUSeaMap can be found in the EUSeaMap 2021 Technical Report . The 2021 technical report describes all modifications and improvements to the methods used to construct the previous version of EUSeaMap (2019) and so users are recommended to also read the EUSeaMap 2019 Technical Report and the EUSeaMap 2016 technical report .
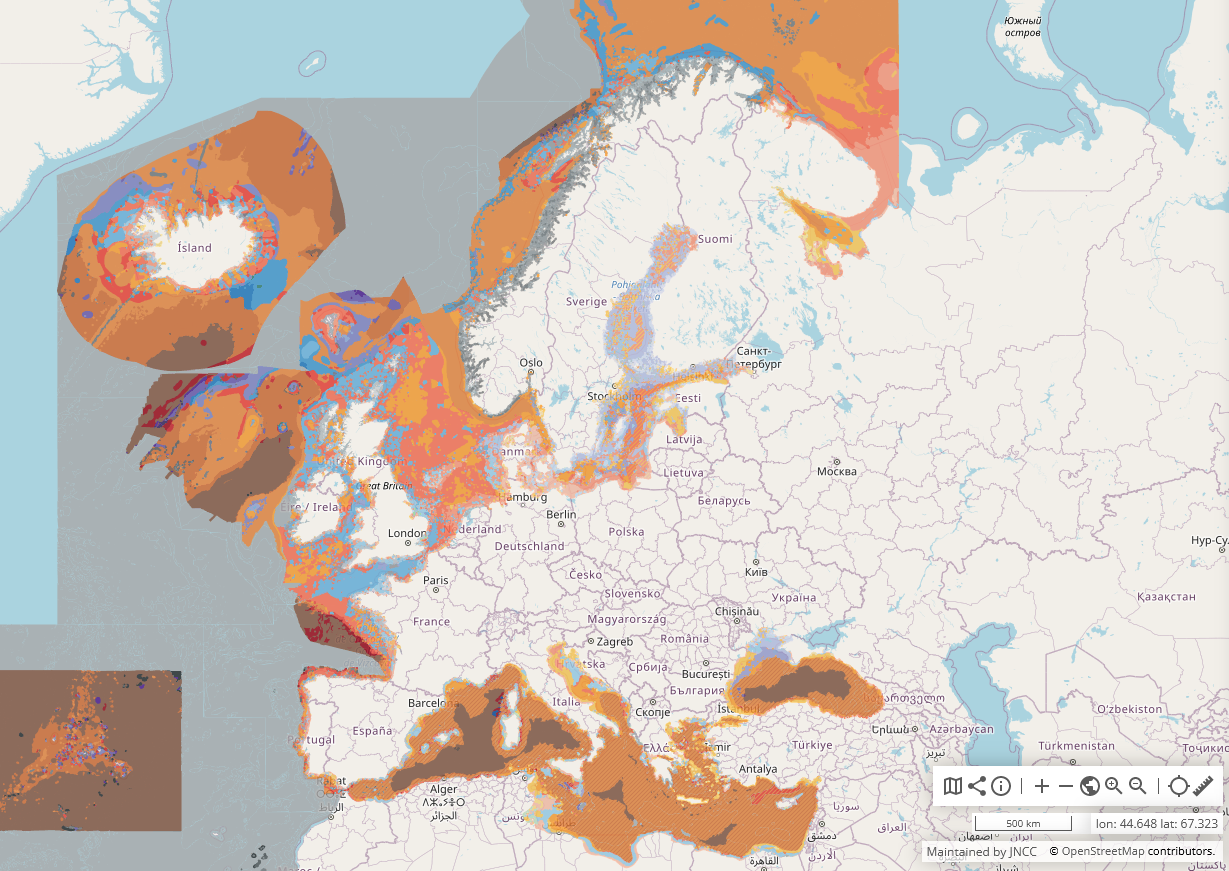
In order to maximise the potential offered by the European marine basins in terms of growth and employment, and to protect the marine environment, we need to know more about the seafloor. European Directives, such as the Marine Strategy Framework Directive (MSFD), call for a full-coverage seabed habitat map of all European seas. In general, habitat maps are very costly and time consuming to produce from survey. The creation of a detailed habitat map involves surveying the seafloor with sonar equipment and collecting samples or photos of the seabed, before analysing and integrating these data types to generate a map. It can take several years from planning a survey to completing a detailed map. By contrast, broad-scale mapping of seabed substrate at a low resolution combined with using modelling techniques to classify habitats in terms of physical parameters is an efficient way to meet the need for a full coverage habitat map at a reasonable cost and a shorter time frame.
EUSeaMap products have been used for assessing and reporting the status of European seas, designing ecologically coherent Marine Protected Area networks, establishing monitoring programmes for seabed habitats and informing marine planning. For more information and some examples of how EUSeaMap products have been applied, see Use Cases.
It is possible to produce a ‘predictive map’ of expected seabed habitat types by combining a series of proxy measurements, such as water depth and light levels amongst others, using statistical analysis and Geographical Information System modelling.
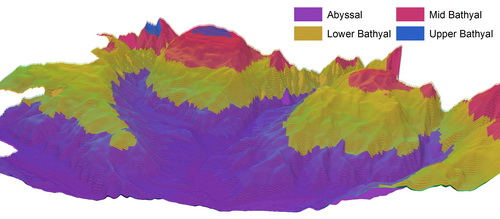
Principal drivers for seabed habitat distributions include the type of seabed substrate (rock, mud, mixed sediment, etc.), depth, light availability and the energy of water movements. To describe the variation in environmental conditions with depth, EUNIS divides subtidal habitats into zones: Infralittoral, Shallow Circalittoral (or Circalittoral), Deep Circalittoral and Deep Sea. In this project we further subdivide the Deep-Sea zone into bathyal and abyssal zones, following scientific literature and recognising the diversity of these huge areas.
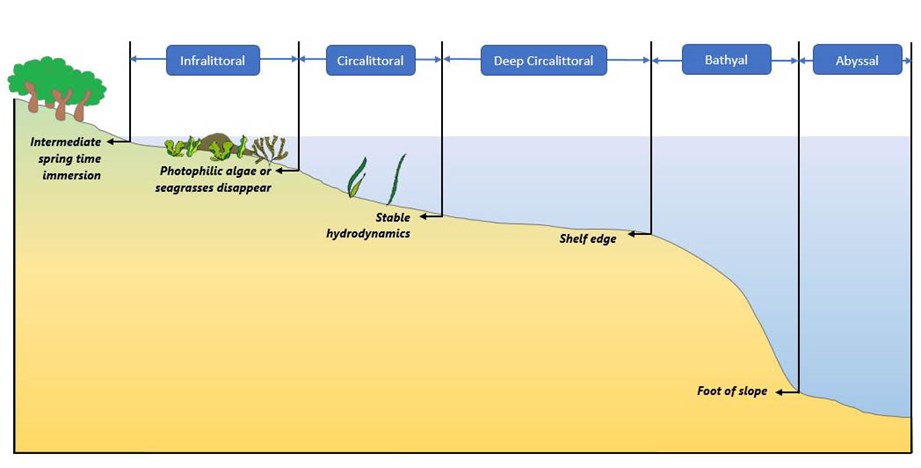
Another factor that can be fundamental in driving habitat types is the degree of exposure to wave and water current energy. For some more enclosed basins other parameters, such as salinity, oxygen levels and temperature of water at the seabed are also considered to be fundamental for habitat mapping.
During the first phase of EMODnet Seabed Habitats (2009-2012), a consistent method was developed to take into account the diverse range of seabed habitats found in different regions.
The first step is to acquire the best-available spatial data for several environmental variables. This includes data provided by EMODnet Geology and EMODnet Bathymetry. The data are organised and harmonised into pixelated images divided into classes. These data can be combined by ‘layering’ the data in GIS to create a combined output describing the habitat.
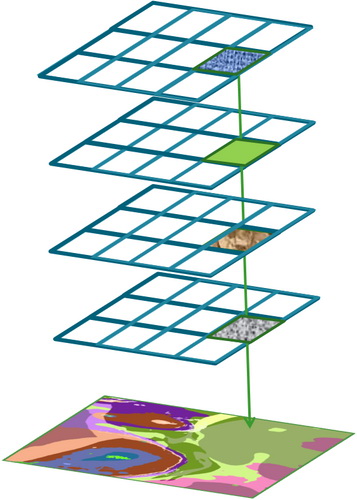
The principal input layers are the type of seabed substrate and the biological zones. Depending on the basin, layers of hydrodynamic energy levels, salinity and/or oxygen are also produced. For example, the hydrodynamic energy layer is divided into ‘Low’, ‘Medium’ and ‘High’ classes.
The EUSeaMap model was developed in ESRITM ArcGIS ModelBuilder and can be saved and executed multiple times, which ensures that the systems are repeatable and easily updated when new layers or methods are available.
In order to create EUSeaMap, it is necessary to define meaningful threshold values for classifying the environmental (input) layers into ecologically-relevant classes for the determination of habitat types. A threshold value indicates where the change in the physical conditions reaches a critical point that defines an expected change in habitat type (at the map-scale adopted in EUSeaMap, 100mx100m blocks). For example, the infralittoral zone is the area of the seabed where photosynthetic algae is able to grow. This is mapped by finding a threshold value of light at the seabed that best fits observations of this limit in the field. More information on EUSeaMap methodology is available in the documents section, including the EUSeaMap 2021 Technical Report , EUSeaMap 2019 Technical Report , and EUSeaMap 2016 technical report .
There have been several iterations of broad-scale seabed habitat maps in Europe in the past decade. The current release of the EMODnet broad-scale seabed habitat map for Europe (EUSeaMap 2021) is the recommended product to use as it supersedes all previous maps (such as EUSeaMap 2019, EUSeaMap 2016, EUSeaMap 2012, BALANCE, MESH broad-scale map, MESH Atlantic).
The latest products are available to view and download for free from the Map Viewer.
-
2023Future Release
- All previously included seas
- European Overseas Territories in the Carribbean
- Caspian Sea
-
2021Official Release
- All previously included seas
-
2019Official Release
- All previously included seas
- Barents Sea
-
2016Official Release
- All previously included seas
-
2015Draft Release
- Azores and Atlantic High Seas
- Black Sea
- Baltic Sea
- Celtic Seas
- Central and Eastern Mediterranean
- Greater North Sea
- Iberia and Biscay
- Norwegian EEZ
-
2014Draft Release
- Adriatic Sea
- Canaries
-
2013Official Release
- Iberia and Biscay
- Azores and Atlantic High Seas
-
2012Official Release
- Celtic Seas
- Greater North Sea
-
2010Official Release
- Baltic Sea
- Western Mediterranean
Data Products Summary
EMODnet Seabed Habitats provides access to a variety of harmonised marine data and products, which are highlighted in the story map and summary tables below. The tables can be used to identify which data and products are available in different basins.
Seabed Habitat Collections
| Product | Atlantic | Arctic | Baltic | Black Sea | Mediterranean Sea | North Sea | Caspian Sea | Caribbean Sea | Other Seas | Total |
|---|---|---|---|---|---|---|---|---|---|---|
| Habitat maps From surveys- EUNIS classification | 321 | 0 | 0 | 9 | 121 | 173 | 0 | 0 | 0 | 559 |
| Habitat maps from surveys - Habitats Directive Annex I | 135 | 1 | 56 | 0 | 90 | 68 | 0 | 0 | 0 | 341 |
| Habitat maps from surveys - other classification systems | 23 | 6 | 7 | 7 | 29 | 46 | 0 | 33 | 0 | 159 |
| Essential fish habitat maps/models | 18 | 8 | 23 | 0 | 6 | 27 | 0 | 0 | 0 | 47 |
| Survey sample points - all | 212,930 | 4,612 | 116,712 | 6,039 | 47,609 | 124,407 | 0 | 0 | 0 | 512,309 |
| Survey samples points - EUNIS classification | 111,643 | 0 | 0 | 79 | 23,249 | 58,224 | 0 | 0 | 0 | 193,195 |
| Modelled maps of specific habitats | 56 | 69 | 47 | 12 | 14 | 63 | 0 | 20 | 22 | 119 |
Seabed Habitat Products
| Product | Atlantic | Arctic | Baltic | Black Sea | Mediterranean Sea | North Sea | Caspian Sea | Caribbean Sea | Other Seas |
|---|---|---|---|---|---|---|---|---|---|
| EMODnet broad-scale seabed habitat map for Europe (EUSeaMap) | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Composite product: OSPAR threatened and/or declining habitats | Yes | Yes | No | No | No | Yes | No | No | No |
| Composite product: Habitats Directive Annex I habitats (external) | Yes | No | Yes | Yes | Yes | Yes | No | No | No |
| Composite product: Essential Ocean Variables | Yes | Yes | Yes | Yes | Yes | Yes | No | Yes | No |
| Composite product: Habitats Directive HELCOM habitats | No | No | Yes | No | No | No | No | No | No |
| Composite product: Biogenic substrate in Europe (draft) | Yes | Yes | Yes | No | Yes | Yes | No | Yes | No |
| Composite product: Coralligenous and other calcareous bioconcretions in the Mediterranean | No | No | No | No | Yes | No | No | No | No |
Physics Products
| Product | Atlantic | Arctic | Baltic | Black Sea | Mediterranean Sea | North Sea | Caspian Sea | Caribbean Sea | Other Seas |
|---|---|---|---|---|---|---|---|---|---|
| Depth to Seabed | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Optical Properties | Yes | Yes | Yes | Yes | Yes | Yes | Yes | No | Yes |
| Salinity | No | Yes | Yes | No | No | Yes | Yes | No | Yes |
| Waves | Yes | Yes | Yes | Yes | Yes | Yes | Yes | No | Yes |
| Currents | Yes | Yes | Yes | Yes | Yes | Yes | Yes | No | Yes |
| Ice Cover | Yes | Yes | Yes | No | No | No | No | No | No |
Chemistry Products
| Product | Atlantic | Arctic | Baltic | Black Sea | Mediterranean Sea | North Sea | Caspian Sea | Caribbean Sea | Other Seas |
|---|---|---|---|---|---|---|---|---|---|
| Density of dissolved oxygen at the seabed | No | No | No | Yes | No | No | No | No | No |
Glossary
A guide to terms and phrases used within Seabed Habitats: Seabed Habitats - Glossary. You may use the links below to jump to a specific bookmark in the glossary.
A, B, C, D, E, F, G, H, I, J, K, L, M, N, O, P, Q, R, S, T, U, V, W, X, Y, Z.
Communication
In this section you will find thematic-specific communication material, and a direct link to the video gallery.